The phrase in the title gives a rather new language in the sciences of human origins. So far the population genetics
(popgenetics) holds opinion that haplogroup R1a appeared somewhere in the southern
steppes, in areas of the modern Russia and Ukraine, according to some 15,000 ybp,
to others 10 thousand ybp, and it waited out Ice Age in the “Ukrainian refuge”. Both
figures came out from the hands of Spencer Wells without any explanation, and without explanation
they were left
when for whatever reason he then changed the number 15 thousand years to 10 thousand years, and how
R1a could survive “Ice Age” in the Ukraine 10 thousand ybp, when the ice age in those days
was
long gone. But the population geneticists continue to preach about the ice age, and about the “Ukrainian haplogroup” R1a, and about 15 thousand ybp. None of them provide a source for
the dating, and no such calculations for the “southern steppes” are given by neither Wells nor
by the others. This
is a normal course for the popgenetics.
At the same time, much data points to the South Siberia and Altai region as the habitat of the
ancient Caucasoids. That was shown by the old works on the analysis of the “genetic distances” in the polymorphic
protein forms (series of works by Nazarov A.F. in the 1980s and 1990s), and b y the research on migration of
haplogroup Q carriers from the Southern Siberia, and apparently specifically from the Altai region, across
the Bering bridge to America. Since the arrival of haplogroup Q carriers to America is dated by
various sources from 20 to 12 thousand ybp, they would have to leave the South Siberia no
later than 40-30 thousand ybp (usually the speed of ancient migrations is assumed to be 1 km per year).
It is estimated that haplogroup Q formed from the combined haplogroup P in the range of 45-40
thousand ybp. Then, the haplogroup P had to be in the South Siberia in those days, and later
form the haplogroups R, R1, R1a, and R1b. All of these haplogroups are anthropologically Caucasoid, and
they came to Europe as Caucasoids, R1a about 11-9 thousand ybp to the Balkans, R1b about 10
thousand ybp to the E.European Plain, and about 5000 ybp to the western and central Europe
(and to the Balkans, to the south-eastern Europe).
1511
This would seem to be a paradox: Caucasoids (Europoids) received their name from Europe, where they came after long
and faraway migrations from the Central Asia. But in fact, it makes sense if we accept possibility that Caucasoids
in fact formed in Europe as a combined haplogroup FT, which spun out onto two branches, with IJK remaining in Europe, and NO + P
migrating to the east, to the southern Siberia, where Haplogroup P rermained for long 20,000 years before
the beginning of its return to Europe (as descendents of the haplogroups R1a and R1b).
Such long-range migrations in academic circles of history and
linguistic are traditionally viewed with suspicion. They just do not have a methodological apparatus
to study such long migrations - many
thousands of miles and for thousands of years, sometimes tens of thousands of years. Usually, the
similarity of material traits at different ends of the continents, and even on different
continents, is held as a chance coincidence. I have numerously read such arguments, for example, as the swastika
sign found at the ancient Indians, where were no Aryans, that that is a chance
coincidence, the same representation of a solar symbol. Frankly, that argument never convinced me,
but I had no objective data to refute it. Now, it have come about in a form of practically identical
ceramics with dozens of identical traits at the Caucasoids (maybe Europeans?)
(and Aryans) and at the American Indians (personal communication of E. Mironov), but that is another story.
The DNA genealogy provides new and compelling evidence of such far-fetched migrations. The
haplogroup R1a has been found in northern China, where its share reaches 30% at a number of local
nations and ethnic groups. But a major issue is not the numbers, but the fact that they are so
riddled with mutations as few (or none at all) haplotypes among other haplogroups. In the northern
China, only short haplotypes of the R1a are available, but the property of the short haplotypes is
that they usually are much more stable than the long specimens. The accuracy of dating is less for
them, but with these “global” mutations, the accuracy is not so important, very important is the
phenomenon itself. The North China haplotypes gave the dating of 20,000 years to a common ancestor
of the R1a haplogroup in that region.
1512
The base haplotype of their 5-marker haplotypes
13 X 14 X X X X 12 X 13 X 30
had only one characteristic difference from the R1a haplotype of the E.European Plain - a third marker (DYS19)
of the latter is usually 16 or 17, and in China it is 14.
The
Altai's R1a haplotypes were investigated in our paper (Klyosov and Rozhanskii, 2012), and it was found that
their common ancestor lived quite recently in historical terms - only 825±320 ybp, and its
core haplotype was
13 26 16 11 11 17 12 11 14 X X 31
It was noticeably different from the E.European Plain base haplotype R1a, with a pair DYS385 =
11-17, which in the E.European Plain is 11-14
13 26 16 11 11 14 12 12 10 13 11 30
A common ancestor of these two basic haplotypes lived 8100
ybp (Klyosov and Rozhanskii, 2012). On the whole, a juxtaposed comparison of the different Altaic
populations' base haplotypes has led to the calculated time to a common ancestor of the European and Altaic
haplotypes for the R1a group in the range of 10,400 and 7,300 ybp (ibid). The same pattern was obtained for
the Tuvan haplotypes; on the whole, a common ancestor of the European, Altai, and Tuvan haplotypes lived
10,000-10,400 ybp (Klyosov and Rozhanskii, 2012).
It should be noted that explorations of the population geneticists are distinguished by the same
flaw, constantly repeated in their “academic” publications. Specifically, it is a static picture and
a lack
of historical comprehension. If they see an R1a haplotype group in Asia, it would “come from Europe”,
it would be immediately called “Scythian”, “Kurgan culture”, “Indo-European”,
without a
slightest justification. In other words, these are “judgments by concepts”, and not by the science. Not
by the science
because they can not calculate dating, and do not include dating in their considerations-reasonings.
In reality, the data and calculations show that in the Altai region, China, Mongolia, South Siberia,
and Central Asia (these regions overlap) are present at least two groups of the R1a haplotype.
One group is the original, the oldest in respect to their common ancestors, not a European, and not
an “Indo-European”, not an Aryan, but strictly autochthonous.
That group is fragmented, it passed various population bottlenecks, and now is constituted of the descendants of
the ancestral lines with age from 600-800 ybp to 7,000 ybp (Mesolithic),
and in rare cases up to 20 thousand
ybp (Upper Paleolithic), as is for the series of the Northern Chinese haplotypes. The haplotypes
of these series are markedly
different from the haplotypes of the E.European Plain.
1513
The other group are haplotypes that really “came” from the Europe 4000-3000 ybp, with their
usually
Aryan, “Indo-European” carriers of R1a. If they can be typed by the R1a subclades, the
result would likely show
the south-eastern subclade L342.2 (R1a1a1h1-L342.2, Middle East, India, eastern part of the E.European Plain, hence South-Eastern
branch) and its derivatives.
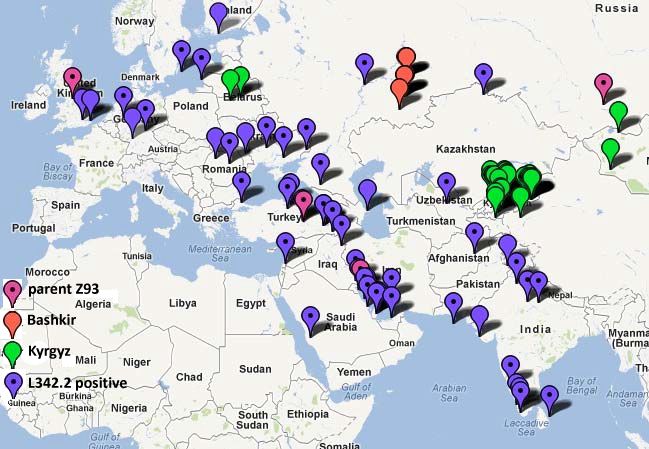
Map 3 (Klyosov and Rozhanskii, 2012)
Map of South-Eastern branch, with birthplaces of furthermost ancestors recorded in available
databases.
Magenta pins denote a parent Z93 branch
red and green pins correspond to Bashkir and Kyrgyz branches, respectively. The rest of the branch
(L342.2 positive) is deep-blue.
The most represented Ashkenazy Jewish branch (not shown) covers densely an area of historical
Polish-Lithuanian Commonwealth.
Therefore for example the Tarim Basin mummies, with the date 4000 ybp, could be either
autochthonous or “Indo-European”, and no rush is in order to call them “European”, “Scythians”, or “Kurganians”.
As has been stated above, for that is no justification. The haplotypes would help, but unfortunately they
either were not defined or not published. The argument that “they wore the clothes like
Scottish plaid” also are not viable, because for consistency it needs to be shown that this type of
clothing was not, oppositely, brought to Europe from the Altai. Consideration of such alternatives is
not innate to the population geneticists.
Archeology is not helpful in resolution of this problem. A study of the Afanasiev culture would probably clarify
the problem, at least partially, but it turned out that for the Afanasiev culture objective dating
is almost nonexistent,
and where its people came from remains unexplored, they either are autochthonous, or the “Europeans”.
The analyses
of the fossil bone remains in respect to the haplogroups were not done yet.
The usual bewilderment about the relationship of the haplogroup R1a origin, essentially Caucasian, with
the Altai, boils down to the fact that the R1a carriers in the Altai (and surrounding areas) are now typical
Mongoloids and speak Turkic languages. The usually asked question is, what kind of the “Indo-Europeans”
are they when they are the Turkic-lingual Mongoloids? That can again be responded with the comment above about the lack of
a historical vision, of the type “what I see, that I sing”. The haplogroup R1a in the Altai, as
is reported above, passed a population bottleneck only 800-900 ybp. What does that mean? That means that these
are not the
Caucasoid ancestors who lived there thousands years ago, although the haplogroup has preserved, at least
partially. This means that 800 - 900 years ago (or at the time) that population was
nearly wiped out, either as a result of natural cataclysms, or most likely by the the enemy.
By their genotype, and hence the phenotype, the surviving R1a carriers and their descendants were already
different people. The fact that they switched to the Turkic language shows who could have been their enemies.
Although the cause could also have been the Turkic-speaking mistresses and wives. If the cause was
the enemies - timewise they
could be Chinese or Mongolian invasions of the 11-13 cc AD. The haplogroup R1a remained at many descendants,
and thus endured at (not numerous) surviving ancestors, but their wives were already not Caucasoid, but Mongoloid.
|
We know of well-documented historical events that illuminate the demographical changes leading to
and in the early 1st mill. AD. These are consecutive relocations of the Türkic nomadic states of
Oguzes (740), Kangars/Bechens (colloquial Pechenegs, 750), Karluks (colloquial Karakhanids, 920),
Kimaks/Kipchaks (colloquial Polovetses, 990) from the greater Altai area to the west. Assuming that
2/3 of the population have relocated, 1/3 of the population minus military and hardship losses
stayed put, and carried on their genotype. The new demographic status at the destinations was a blend of the previous
population and the newcomers from the immediate east, linguistically also Türkic, but with
relatively higher proportion of the eastern Eurasian Y-DNA and mtDNA genes. The new geographical layout remained relatively stable for 200 years, until the turmoil of the
Chingizkhan. Chingizkhan subjugated and wiped out the surrounding linguistically Türkic and Mongolic
people, but his wipeout achievement zeroed on the Mongolic division of the three-division Tatars,
leaving the Uigur division intact, and also leaving intact the nominally Tatar component of the
Kimak Kaganate. Chingizkhan was a recognized offspring of the Uigur Kaganate dynastic line Jalair,
via a Borjigin ancestor, and supposedly belonged to the Uigur Jalair Y-DNA line. As a result, after
the “Tatar wipe-out”, the subsequent movement headed by few thousand Mongols became known as a Tatar
movement.
The Tatar movement, demographically nearly exclusively linguistically Türkic, brought about
further demographic outflows from the greater Altai area. Assuming that 2/3 of the population have
relocated westward, the 1/3 of the population minus military and hardship losses stayed put, and
carried on their genotype.
That is “the enemies - timewise they could be Chinese or Mongolian invasions of the 11-13 cc.
AD”. There were no Chinese or Mongolian invasions of the 11-13 cc. AD, the Chinese were cornered and
could not fend for themselves, and Mongolian nomadic drifters were just filling in the voids left by
the “Tatar” migrations. On the other end of the Urals, at that period occurred a corresponding
demographic change with the opposite effect, in the Eastern Europe formed and endured a new numerous
nation that absorbed predominantly Türkic existing populations and the new Türkic migrants from the
east, known as Kipchak Khanate and Tatar Khanate (colloquial Golden Horde, 1240).
Demographically, few of the continuous 6-centuries long Altai migrations to the Eastern Europe
reverted back to their greater Altai domicile. The majority were absorbed by the new powers-to-be,
adding their genetic markers to the local ethnicities. It is inconceivable that the genetic markers
of the early Altaians would not be found in the Eastern Europe and in the Balkans; the genetic
markers remaining in situ in the C.Asia had to be subjected to the stresses and dilutions of the coming centuries.
By the early 13th c., of the mid-1st mill. AD population, in the Altai probably remained only 10%
(1/32) of the early Altaian population, 90% of the early Altaians having moved to the
Middle Asia and Eastern Europe. At least partially, the outflow was replenished by the kindred genotypes migrating from the east.
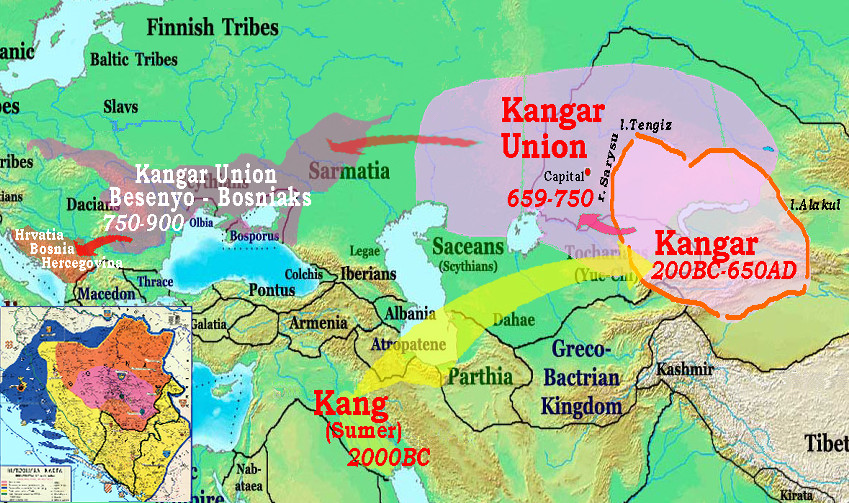
And then in 1650 fell a Jungar genocide. Numerous populations can be traced individually, for example the Kangar anabasis below:
The influence of the female Mongoloid genotype can't be underestimated, and should not be
overestimated. The Mongoloid admixture, also termed Lapanoid and Uralic, is traced in the eastern and central
European population from the Paleolithic times; every study of the Kurgan people and steppe nomads from the later Mesolithic times to the
Middle Ages noted their Caucasoid/Mongoloid admixture, and notoriously more notable Mongoloid appearance of the
females than of the males. To suggest a supposed linguistic replacement to the effects of the
Türkic-speaking mistresses and wives mass-deranging the noble IE speakers is an open-ended
proposition that affects all ages down to the Paleolithic times. A constant accumulated increase of
the Mongoloid component is observed in the Middle Asia/S.Siberia from the 7th c. BC.
For horse-mounted nomads, the greatest influential demographical factors are migration (exodus),
followed by genocides, followed by exogamic law. |
1514
That is precisely what changed the genotype and phenotype of the offsprings while preserving the male
chromosome and its haplogroup R1a. Therefore, can be expected a shift of the mtDNA spectrum toward
the Asian Mongoloid types of the
mitochondrial haplogroups. As is seen below, this is how it turned out.
It is clear that with such state of the matter, the new scientific publications, with new material, and
with substantiated conclusions are of undoubted interest and importance. And in this 2012 year came
out two
articles on the “Altai subject”. The first is a joint article of the specialists from the Penn
university (Matthew Dulik, Theodore Schurr, et al.), from the Institute of Cytology and Genetics of
the Russian Academy of Sciences Siberian Branch (Sergey Jadanov and Ludmila Osipova), and from the
Kazakhstan Institute of General Genetics and Cytology (Aiken Askapuli), entitled “Mitochondrial DNA and Y Chromosome
Variation Provides Evidence for a Recent Common Ancestry between Native Americans and Indigenous
Altaians”. The second is the article by Spanish authors (Barcelona) Gonzales-Ruiz, Malgosa, et
al. “Tracing the origin of the east-west
population admixture in the Altai Region (Central Asia)”.
As usual, hopes for the emergence of advanced works were justified only partially. Both works contain
interesting findings, which however are largely drowned in a swamp of incongruities. More interesting
turned out to be the first work, but it did not answer our questions outlined above. Nevertheless,
it contained raw material
that enabled to do some (and very informative) calculations shown below.
Let's start with the second, much simplistic work (in the context outlined above).
Article by Gonzalez-Ruiz et al. (2012)
Gonzales-Ruiz and coauthors investigated mtDNA in the skeletal remains of the Pazyryk culture. This
is an archaeological culture of the Iron Age (3rd - 5th centuries BC, although some extend the date to the
6th century BC) (see Marsadolov L.S., 1994, Pazyryk Timing, and
A.Yu.Alekseev et. al., 2001, Chronology of Eurasian Scythian Antiquities
Born by New Archaeological and 14C Data), which is
(archeologically) classed as belonging to the “Eastern Scythian circle”.
The area is the Mountain Altai and
adjacent areas of the Altai, Kazakhstan, and Mongolia. Economic mainstay is nomadic animal breeding. It is
suggested that Pazyryk is a derivative of the Afanasiev culture. Its main physical types are dolichocranic Caucasoid
with high and wide face (probably, carriers of haplogroup R1a), brachycranic Mongoloid with low face, and mezzo dolichocranic Mongoloid
with medium height face (the latter can be carriers of mtDNA C, N, Q, and their paired haplogroups).
1515
It should be stated that the previous paragraph in the article is hardly present. Instead, the authors
expand that the Altai is a boundary of the western and eastern Eurasian steppes, especially dwell on the
“Scythians”, who were “Caucasoid” (called “Europoid”) and
spoke “Indo-European languages”. Altrogether, the standard
bouquet of population genetic doctrines, including mentioning of Herodotus. No other data that the
fossil remains the authors studied belonged to the Scythians are stated in the article, and it is unclear why
it was needed at all to write about the
Scythians. It is not known, whether these are Scythians. Right there goes on the “nomadic pastoralism”, although bones also do not tell about it. In short, flows a
powerful that to the material content has no relation (probably, the
background summarizes contents and terminology of the bibliography). But this is a norm in popgenetics.
That in the tombs were found Caucasoid anthropological features is immediately presented as “the Scythians
had a European morphology”. That Caucasoid morphology could remain of the the indigenous Caucasoids had not occurred
to the authors. This is the popgenetics.
The authors examined mtDNA (HVRI, hypervariable region) of 19 skeletal remains from four tombs in the Mongolian
Altai, dated to the Bronze (three mtDNA) (3000-700BC) and Iron (16 mtDNA)
(800-300BC) Ages.
|
Studies of pre-historical periods without reasonably accurate dating is senseless, and any
attributions and conclusions incredible. |
It was postulated (more accurately, accepted as given) that the mtDNA haplogroups are
divided into three regional sources:
- West Eurasian: R0 - R0a'b, HV, N1, JT, UK, W, X
- East Eurasian: M - C, D, G, Z, M9, M10, M11, M13, A, B, F, N9a
- South Asian: M*, U1a-c, U9, R*, R1-R2, R5-R6, N1d.
Then went typical popgenetic correlations, like the “diversity of genes”, “nucleotide
diversity”, Harlequin, FST, “paired genetic distances between populations”, and so on,
that in the DNA genealogy do not make sense, but there is some mysterious meaning in popgenetics,
which is OK. Instead of relaying
that alchemy, let's look at once what is it in essence. That would be constructive.
So, of the 19 remains 11 turned out to be haplogroups A, C, D and G, i.e. the “East Eurasian”, 9
turned out to be J, K, HV,
U, and T, i.e. “West Eurasian”. The authors chose not to provide these numbers directly,
9 and 11, but show “58%” and “42%”, otherwise the quantities are too small.
In reality, of the first 11 haplogroups seven turned out to be haplogroup D - all three haplogroups
of the “Bronze Age”,
and the other four of the Iron Age. The A, C, and G were one each, all of the Iron Age.
1516
In the Iron Age Altay sample were found three K, two U (U5a1), and one J, HV, and T each. This
allowed the authors to conclude that in the Bronze Age all (three) haplogroups were “oriental”, and
in the Iron Age they were complemented by various “Western” haplogroups. That is the “admixture”.
Loosely speaking, that may be right.
However, from the Bronze Age came only three samples, and from the Iron Age came 16. Curiously, what would
be the distribution, if it was the opposite, 16 of the Bronze Age, and 3 of the Iron Age? Obviously,
the distribution would be different. However, the authors tagged the Pazyryk population “genetically homogeneous”.
Taking a picture as given, it comes out that the brides and wives of the Altaians (mostly R1a, as
is shown below) were East Asian women (probably Mongoloid), and in the Iron Age were added Western
Eurasian women. Either they were coming themselves, or they were abducted.
That is, in essence, the entire article. Everything else is general chat. Certainly, from the
article is not much to learn. Although the fact of the analysis of the fossil mtDNA is a no small achievement
that requires some technical scientific school.
Article by Dulik et al. (2012)
As was mentioned above, the article by Dulik et al. is much more advanced.
True, the main issue she addresses, namely the origin of the Amerindians, with a
conclusion that originally they are from the Southern Siberia, has been settled in the literature long
ago. And that they have common haplogroups Q and C has long been resolved too. The authors delve into
details of exactly which subclades they have in common, but in my view it is unproductive, other
than as general exercises on the topic. Well, they have found common subclades (actually, missed more
than found), and what it resolves, what problem it answers? Moreover, the authors got jumbled between
the “genealogical” and “population” mutation rates, and eventually came to a totally wrong conclusion,
completely bungling discussion and conclusions.
It is quite regrettable that the authors are completely unfamiliar with the current literature on
the mutation rates. For example, this author three years ago has published a detailed article in the
J. Genet. Geneal. (2009) and Human Genetics (2009) what are the actual constants of the
mutation rates, and how
they are calibrated.
1517
Well, the authors do not read literature on the mutation rates, alas, then why publish
illiterate articles on the subject? That's becides the fact that numerous forums have long debunked the
“Zhivotovsky population rates”, and it seems the authors are unaware of that. How do you
have to disrespect the subject of their own research not to be interested, not to read the
forums, and finally, do to read the journal “Proceedings of the Academy of DNA Genealogy”,
where fifty
issues starting from 2008 sorted out the mutation rate constants dozens of times, repeatedly
demonstrating that the “Zhivotovsky population rates” for the real systems are sheer
nonsense, moreover, essentially they are a product of doctoring and outright manipulation of the
data. On that, Proceedings devoted special articles. Nevertheless, the authors cite Zhivotovsky, although
they seems to have already been aware
that something is wrong there. So they calculate in parallel “populationally” and “genealogically”, which leads to a
complete mess in the results. And so they write accordingly that calculating “populationally” it comes out
recent, but calculating “genealogically”, it comes out as the Bronze Age. Well, this is
not serious. However, in the “popgenetics” anything is possible. Like in shamanism.
But the funniest matter is not even that. In the end, the authors came to a conclusion that the “population rate”
more correctly describes the data, agreeing better with the data of the history. Thus, the “population” rates for the
South American Q1a3a1a* produce the time to the common ancestors (according to the authors) in the
America 4.9 thousand years and 7.7 thousand years, and “according to Zhivotovsky” 22.0 and 13.4 thousand
years, which means that the second is correct.
This, of course, boggles the mind. Have the authors heard about the “population bottlenecks”?
How come they suddenly decided that any population in the calculation gives exactly the “true”
value of the time to the ancient common ancestor? For example, according to the authors, the series
of the Altai Q1a3a* (which should be older by phylogeny than the Q1a3a1a*) of the 19 17-marker
haplotypes has only 29 mutations from the base haplotype
13 24 13 10 15 18 Х Х 13 14 14 17 –
16 14 19 11 16
11 22
and on the top of that, these markers do not have any mutations. It is clear that a common ancestor of these 19 haplotypes
is recent. Indeed, 29/19/0.034 = 45 → 47 conditional generation (at 25 years each), i.e. 1175±250
years to a common ancestor. And by the “logic” of the authors, it should be not less than
20 thousand years. Hence, the “method is wrong”, is it not? But this just means that the ancestors
did not survive in their offsprings, the DNA genealogical line almost cut short, the population has
passed through a bottleneck, or a “genetic
drift” as say the popgeneticists. In short, the common ancestor of the series lived recently, and
there is nothing wrong. This also happened in the America with a series Q1a3a1a* haplotypes. The
series should not be artificially
extended three times using a flawed method.
1518
The “advancement” of the article is not, of course, that calculations were wrong, and that
it “discovered
America”, which has long been discovered. But that the article published a good haplotype
compilation for
different Altaic populations, as many as 189 haplotypes in the 17-marker format. It would
be better have haplotypes been longer, but what has been published is also alright. It would be better if
the subclades of the R1a1a1 were revealed,
but the published is also alright. But they measured only R1a1a-M17 (R1a1a) and R1a1a1-M417 (R1a1a1) [the first SNP
was
not needed, in this situation it is redundant, since all haplotypes had M417], and stopped at that, incorrectly noting
R1a1a1* (with asterisk), as it has no downstream SNPs. That M458
(Central European and West Slavic branches) and L365 (North-European branch) were not found, is
natural, they could not be found. It
is known that there are none in the east, except for the recent “tourists”. But to measure L342.2
would be appropriate, it would separate the autochthonous subclades and the “Indo-European”.
Too bad that
the authors probably did not know of that snip.
The authors found that the studied populations mostly had “East Eurasian mtDNA haplogroups”, which
generally contradict the previous paper cited above, according to which from the Iron Age
began an active “mixing” of mtDNA. In reality, the haplogroups A, C, D, and G turned out
to be in majority at all tested
populations:
- at Chelkans (Chelkandy, N.Altai, a part of Tubalars) (38%), another 19% N9a,
the other are minor;
- at Kumandins (Kumandy, Kumans, N.Altai, with seoks Chabat, So/Solu;
phenotype Ugrian Hanty/Mansi) (62%, the rest are minor);
- at Tubalars (N.Altai, Turgeshes with seoks Chagat/Chygat of Tele stock,
Chalkanyg; Kumysh/Komlyash/Komnosh aka Black Tatars, Mountain/ Taiga Tatars, Kumysh = Silver; Kuzen,
Palan-Komdosh/Bolan, Tiber, Togus/Tokuz = Tokuz-Oguzes, Turgesh/Kergesh, Yalan, Yus, Yus-Shanmai) (64% and 53% for the two populations, the rest are minor);
- at Shors (N.Altai, with seoks Aba/Aban, Chelei/Teleut, Kalar, Chelei/Elei,
total 22 seoks; language of Uigur family; phenotype Ugrian Hanty/Mansi) (50%), another 14% in F1, the other minor;
- at Altai-kiji (N.Altai, with 36 seoks Irkit, Naiman, Kypchak, Mundus/Kypchak
the largest seoks; Tele 30%, Türks/Ashina Türks 22%, Enisei Kyrgyzes 12%, Naimans 10%) (48% and 36% in the two populations, the rest are minor)
- at Telengits (S.Altai, Tele tribes Telengut, Telengit, and Tele) (40%), 15% of haplogroup V, the rest are minor, and
- at Teleuts (S.Altai, Tele tribes Telengut, Telengit, and Tele) (51%, the rest are minor).
This is important, and confirms the above hypothesis that despite the Caucasoid
(Y-DNA) haplogroup
R1a, Mongoloid women turned R1a carriers from Caucasoids to Mongoloids, and even changed their language to
Türkic.
|
In retrospective study of the Türkic tribes' origin, Altai served as a refuge, over millenniums
refugees from many surrounding lands found safety in Altai. The layers of refugees superimposed,
intermixed, and merged like layers of old paint on an antique furniture. Peeling one layer after
another, in many cases the origin of the Altai tribes can be tracked down to their first naming in
the Chinese dynastic chronicles, in the works of antique Greek authors, and other antique sources.
For the Türkic tribes, Altai, like just like Caucasus and Khingan, was a shelter with ongoing
historical merging, and not a source from which emanated numerous Türkic tribes and peoples.
Complimentary to Altai, the Minusinsk depression of the “Karasuk time” in the middle of the 2nd
millennium - 7th century BC was a stopover location in the spread of some Türkic tribes, possibly
together with the Ket tribes. A good case study example is the case of Tokuz-Oguz (Nine tribe confederation, aka High Wheels,
Ch. Dinlin/Dingling) tribes Apa/Atie/Adie, Beiho/Huiho/Husie or Uigurs, Dubo/Tubo, Guligan/Kurykan =
Sakha/Yakut, Kibi/Kipi, Paegu/Baegu, Pugu, Seyanto, and Tunlo, which in the words of Chinese annals
constituted a “special generation of Tele”, and the tribes Fulo/Buri, Fuliku/Buriku, Hun, Husie/Husa,
Sekis (Se/Saka), Taki, Teleer, and Telengit/Tolanko/Dolange, which also were treated as Tele tribes.
It has long been established that Tele tribes were Türkic-speaking. A corollary of that is that
the mixed phenotypically Caucasoid/Mongoloid men with genetically Caucasoid/Mongoloid genes, and the
mixed phenotypically Caucasoid/Mongoloid women with genetically Caucasoid/Mongoloid genes spoke the
Türkic language. What was the language of the female's ancestors, as well as the male contribution
from the eastern lineages, may be clarified one day with the help of the ongoing genetic studies,
but a linguistic mass-conversion of the noble IE speakers, without a grain of detected noble IE
substrate in the body of the 42 mongrel Türkic languages, is a pathetic pipe dream.
Not all of the Tele tribes were nomadic horsebreeders. Not all of the Tele tribes were elite
tribes either. The Chinese annals illuminated the useful, visible, and dangerous, leaving in the
shade non-prominent and dependent kyshtym tribes (kyshtym = allied dependency/tributary). These
tribes economically were at a different level from the horsebreeders, their traditional sedentary
subsistence was in hunting, fishing, farming, small ungulate husbandry, charcoal, metal smelting,
metalwork, and such. At the turn of the eras, numerous Tele tribes and confederations occupied
sub-taiga woodland belt from Khingai in the east to the Urals in the west. Neither Chinese annals,
nor the later writers reported on social relations within the Türkic confederations, but the first
records on the Türkic tribes that fled to Altai in the 16th-17th cc. reported that the elite tribes
were fleeing with their kyshtyms, and the local elite Enisei Kirgiz tribe had numerous kyshtym
dependents across Altai and sub-taiga woodland belt. Understanding the ethnic and social
relations is a necessary condition for genetic sampling and conclusions of the ancient and modern
Altaians.
One of the Tele tribes was Shibir, it was an elite tribe that gave the Türkic Kaganate at least
one Kagan (609-619), probably from the Ashina Kagan clan and Sibir Hatun clan. Shibir tribe is
listed among the Tele tribes that Juchi subdued in 1207, along with the forest tribes of Kesdiin,
Bait, Tuhas, Tenlek, Toeles, Tas, and Bachjigi. Shibir was the westernmost tribe, and the
easternmost tribe Bachjigi was occupying the Sayan-Altai uplands. At about the same time, the tribe
Sabir/Savir/Subar/Suvar/Suvar gained an upper hand and a title “King” of the Armenian chroniclers
over the Kayi and Masgut tribes in the E.Caspian seaboard in the Caucasus. The first records on the
nomadic Sabirs come from the Babylonian cuneiform tablets of 23rd-22nd cc. BC, along with the
Mesopotamian horse nomadic tribes Kuman, Quti, Lulu, and Turuk. Thus, we have literary traces on the
tribe Sabir from the 23rd-22nd cc. BC to modernity, from the Mesopotamia to Khingan, illustrating
the author's point on popgenetic's static picture and a lack of historical comprehension. |
1519
Now turning to the most important for us (in the DNA genealogy) object, the haplotypes. Of the
189 haplotypes 75, i.e. 40%, were R1a, of them the majority were 60 haplotype R1a at the population
of Altai-kiji
(Tele 30%, Türks/Ashina Türks 22%, Enisei Kyrgyzes 12%, Naimans 10%). For
comparison, haplotype R1b was found only 6 ea., or 3%, all in the population of Kumandins
(Cumans/Kumans, Ugrian Hanty/Mansi phenotype, 62% eastern mtDNA). The second
highest number was Haplogroup Q, total 34 haplotypes (18%), five subclades. Haplogroup C (in
two subclades, C3* and С3с1) included 24 haplotypes (13%). Haplogroup N in four subclades (N1*,
N1b*, N1c* and N1c1 [last only two haplotypes]) included 22 haplotypes (12%). The remaining
haplogroups - D3a (six haplotypes), J2a (three haplotypes), E1b1b1c, I2a and L (one haplotype). This
is all, so to say, again popgenetics, and refers to what we have NOW. To the DNA genealogy
it has almost no relationship. All these individual haplotypes from the 189 as a rule are accidental, for
example from the groups I2a, E1b, J2a in southern Siberia. What is no accident, it is the 75 haplotypes
of the Caucasoid
(originally) haplogroup R1a, and if in the west they are mainly mtDNA of the group H, in the Altai
they are mostly mtDNA of the group C. There lies the reason for their Mongoloid phenotype.
Altai-kiji (Tele 30%, Türks/Ashina Türks 22%, Enisei Kyrgyzes
12%)
To put things in perspective, here is an example of 1897 census, totaling 19,000 Altai-kiji people.
The compatible number for 2002 census was 62,000. The rate of population growth points to significant dilution
of the 1897 genetic inventory. But the number of the1897 census, taken by taxing
district (volost, a distorted Türkic ulus with a prosthetic v) with mixed
population, was already extremely mixed, with the Altai-kiji proper constituting only 8% (1520 total)
of the listed Altai-kiji. A blind sampling of the modern Altai-kiji is ethnically as informative as
testing for its origins a herring gumbo soup (data from
L.P.Potapov, 1969, “Ethnic composition and origin of Altaians”, Science, Leningrad).
| 1 |
Todosh |
2978 persons |
19 |
Merkut |
166 persons |
| 2 |
Irkit |
1928 |
20 |
Koojo |
139 |
| 3 |
Naiman |
1912 |
21 |
Kaan |
118 |
| 4 |
Kypchak |
1681 |
22 |
Almat |
105 |
| 5 |
Mundus |
1342 |
23 |
Chagandyk |
89 |
| 6 |
Teles |
1333 |
24 |
Modor |
72 |
| 7 |
Kergil (Turgesh) |
1035 |
25 |
Tumat |
72 |
| 8 |
Tongjoan |
974 |
26 |
Derbet |
56 |
| 9 |
Soen |
684 |
27 |
Bogus-han |
49 |
| 10 |
Kobek |
565 |
28 |
Yaryk |
28 |
| 11 |
Chajats |
558 |
29 |
Yus |
19 |
| 12 |
Ochy |
528 |
30 |
Saal |
15 |
| 13 |
Bailagas |
398 |
31 |
Purut |
9 |
| 14 |
Koobaly |
348 |
32 |
Elik |
7 |
| 15 |
Dieti-Sary |
295 |
33 |
Mogol |
3 |
| 16 |
Olun |
275 |
34 |
Meret |
2 |
| 17 |
(garbled) |
245 |
35 |
Togus |
1 |
| 18 |
Gandi |
229 |
36 |
Suuzar |
1 |
| Total |
19,000 |
|
|
|
|
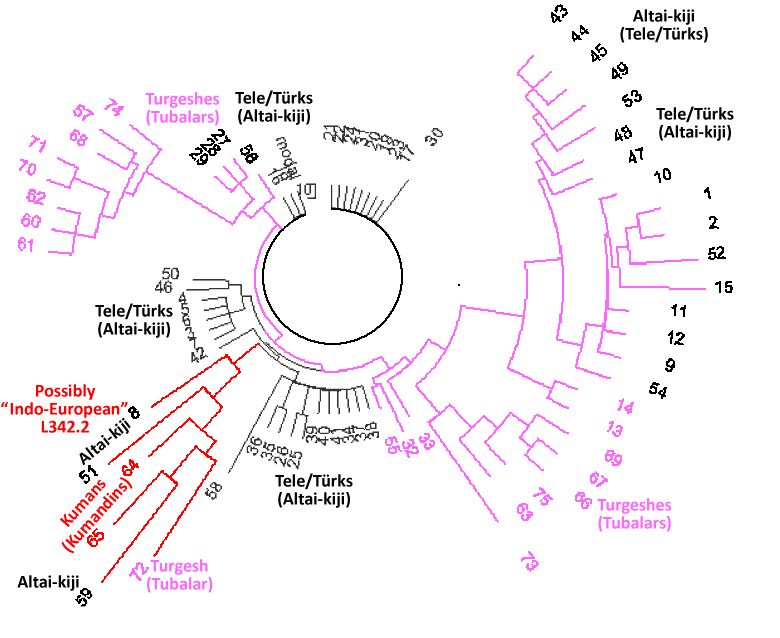
Fig. 1. The tree of the 75 haplotypes of haplogroup R1a1a1-M417 in the Altai.
Upper left arm - Tubalars (57, 68, 70, 71, 74), Chelkans (60, 61, 62) and Altai-kiji (57). The
lower right-hand branch - similar composition: Tubalars (66, 67, 69, 73, 75) and one Chelkan (63).
The lower left branch - mixed: Kumandins (64, 65), Tubalars (72), Altai-kiji (8, 51, 59). The
remaining haplotypes - Altai-kiji.
(color coding added, ethnic affiliation conditional to statistical
probability of fraction in population and randomness of sampling)
1520
Figure 1 shows the tree of 75 haplotypes of haplogroup R1a in Altai.
One can see that the tree has several completely different branches. For a single haplogroup, so unusual tree is rare, and usually indicates a series of population bottlenecks, which split onto branches
in early history, and has barely survived in relatively recent times. The massive differences between
the branches indicate that the original common ancestors belong to the far antiquity.
Mixing up and averaging all haplotypes of the tree, which would be totally wrong, would generate
a phantom “common ancestor”
who ostensibly lived about 4,300 ybp. To do that is wrong because the tree consists of branches, each
one of
which describes a separate DNA-genealogical line, and the branches are of different size, and
correspondingly of different “weight”. With averaging, the most ancient branches are lost, and the tree is
“rejuvenated”. In
the “Zhivotovsky method” the rejuvenation is artificially by increasing the age by about three times.
Why specifically three? So yes. So showed the theory for 10,000 haplotypes with their full homogenization
for the
history of the whole humanity, which is totally artificial and unrealistic premise. And the “compensation”
is most “dull”, it presumes that correction by a factor of three is independent of the haplotype
format, whether it is 6-marker, 9-marker, 12-marker or 17-marker. Or any other formats. That is, it is
presumed that the mutation rate is
independent of what the haplotype markers are and how many are there. It is inconceivable that none of
the “population geneticists” pay any attention to that. At least someone should have thought that this simply can
not exist. That's what a lack of adequate scientific school is. A complete atrophy of thinking. In
short, using the “Zhivotovsky method” it comes out that the time to the common ancestor of the
whole tree is 12.800 years. In reality, this answer is not even close.
Let's consider the haplotype tree Fig. 1 in terms of the DNA genealogy.
Kangars/Turgeshes, modern Tubalars and Chelkans
The eight haplotypes in the upper left branch have a base haplotype (in the format of Y-filer,
i.e. DYS
393, 390, 19, 391, 385a, 385b, 439, 389-1, 392, 389-2 - 458, 437, 448, GATAH4 , 456, 438, 635):
13 24 16 9
12 14 10 14 11 18
– 14 14 20 12 17
11 23 (No 57, 60-62, 68, 70, 71, 74)
1521
The branch has 10 mutations (in the highlighted markers;
the other markers have no mutations), which
gives 10/8/0.034 = 37 → 38 conditional generations, or 950±315 years to a common ancestor
(the arrow indicated a correction for reverse mutation, 0.034 is a mutation rate constant for 17-marker
haplotypes). This haplotype is notably different from the base haplotype in the E.European Plain (by 9
mutations)
13 25 16 11 11 14 10 13 11 17 - 15 14 20 11 16 11 23
and from the base southeast haplotype subclade L342.2 (by 8 mutations)
13 25 16 11 11 14 10 13 11 17 - 15 14 20 12 16 11 23
That places their common ancestor at 7250 ybp (6750 ybp
to the common ancestor with the subclade L342.2). This could not be the “Indo-European” R1a, in those days they
(the R1a “Indo-Europeans”) lived in Europe, and had not even
arrived yet in the E.European Plain. This is an autochthonous common ancestor, most likely a
phantom ancestor (i.e., the dating can be significantly underestimated), but this is not an “Indo-European” common
ancestor. The modern fairly young population consists mainly of Tubalars and Chelkans
(Turgeshes). Now they are Mongoloid and Türkic.
|
Tubalars and Chelkans are descendents of the Turgesh tribe, aka Kengeres and Kergej, supposedly a
descendent of the Kangar/Kangju state. In 651 -766 Turgeshes established their own Kaganate, they
suffered great losses in the wars of independence against encroaching China and pretentions of the Türkic
Kaganates, they carried a brunt of the Arab conquest campaigns in 711 - 724, suffered from
internecine wars of succession, and finally in 766 were dismembered by Karluks (Uigurs), who
displaced them from the Jeti-su area. During their independence, Turgeshes suffered numerous
genocides, and vanished as a great power. A part of the Kangars/Kangly retained their state, known
as Kangar Union (659-750) northwest of the Jeti-su, but eventually had to escape to the N.Pontic
(under the name of Bechens/Bajanaks/Pechenegs, 750-990), and then to Balkans, establishing Hrvatia/Croatia
and Bosnia-Hercegovina. A second recent bottleneck is connected with the 13th c. Mongol expansion, when numerous Türkic
tribes under Mongol flag campaigned and settled in the China and E.Europe, considerably reducing
their presence in the Altrai, S.Siberia, and Middle Asia. Genetically, it is reflected in replacement
among their descendants in the E.Europe of the 2/3 of the eastern mtDNA with the western mtDNA
haplogroups.
The last bottleneck is connected with the Teleut Tele tribes escaping to the then-Russian Altai
fleeing form the army of the Manchurian-ruled Chinese Empire Qing exterminating Jungars and their
former kishtyms without any exceptions (events of 1750s - 1790s), bringing nearly complete extermination
of the Tele people, when no more than 10% of the Tele-confederated people survived
and found refuge in the mountainous areas in the Altai, Sayans, Pamir, and Chinese Turkestan.
Due to the pinpoint accuracy achieved by the DNA genealogy, we have a chance to peek into the
male and female genetic composition of the Kangars/Turgeshes, and trace their lineage in the modern
diaspora. The supposition that Kangars/Turgeshes populated Altai before or after displacement of
population in the Altai is unjustified, the Kangar phenotype and ontological descriptions, their
Jetyasar archeological culture, their kurgan burial customs, their popular cranial deformation
traditions, and even their coinage are detailed in the literature and can be traced across Eurasia.
The Altai findings reflect not only a different epoch, they also belong to the different
geographical area as much as the DNA of the modern Caucasoid Australians describes DNA of a different
geographical area.
|
Potentially Türks/Ashina Türks, or their parent tribe of Saka,
a part of modern Altai-kiji
At the foot of the described branch is shown a mini-branch of four haplotypes, three of which are
identical, and the fourth (56) differs by only one mutation (in DYS390).
13 26
16 10
11 14 10 14
11 18
– 15 14 21 12
16 11 23 (No 27, 28, 29)
13 25 16 10 11 14 10 14 11 18 – 15 14 21 12 16 11 23 (No 56)
|
Of these 4 branches, 3 may belong to the second-largest component of the Altai-kiji, the Türks/Ashina
Türks 22%, or their parent tribe of Saka |
Six mutations (marked) separate a common ancestors of these haplotypes and the basic haplotype of
the E.European Plains by 6/0.034 = 176 → 214 generations, or 5,350 years, and places their common ancestor
by (5350+4600)/2 = 4975 ybp. It is also too early for the arrival of the “Indo-Europeans” R1a to the Altai. They
(the “Indo-Europeans”) arrived a
thousand years later (“Indo-Europeans” in quotation marks are neither Indo-
nor Europeans, the label comes from the attribution by the latter days seers of the Andronovo Kurgan
culture to the Ossetians aka Iranians aka Indo-Europeans aka most reverend ourselves).
One mutation between the above two haplotypes places the mini-branch at 1/0.34 = 29 → 30 generations, or
about 750 ybp. But between this mini-branch (Altai-kiji) and the branch of the Tubalars and Chelkans
(Turgesh branch)
are
6.5 mutations, or 5900 years between their common ancestors. This puts THEIR common ancestor at
(5900 + 950 + 750) / 2 = 3800 ybp.
|
The indiscriminate blend of the Tele and Saka within the Altai-kiji blend, and the Kangars/Turgeshes
genetically separated 3800 ybp, at about the time when the E.European agricultural refugees
just started their march southeast toward the Indian peninsula, and the Zhou Kurganians of South
Siberia reached the prehistoric China. |
In other words, from the DNA line running from 3800 ybp (which in turn also clearly
passed the population bottleneck) remained two sub-branches, aged 950 and 750 ybp. But the distance between
them shows that from their common ancestor they separated far apart, and that distance can be easily
calculated.
1522
Thus, we see that the upper left part of the tree is not
an “Indo-European” branch, but autochthonous haplotypes
of R1a. They are ancient in origin, much older than the haplotype R1a (more precisely, their common
ancestors) in the E.European Plain.
Teleut Tele
This type of calculations allows to obtain the following basic haplotypes and ages of
the common ancestors of the branches. A small branch on the left of seven haplotypes (with total of
only three mutations)
13 25 16 11 11 14 10 14 11 18 – 15 14 21 10 16 11 23 (325 years to a common ancestor)
(No 3 - 7, 42, 46, 50).
|
325 years nearly agrees with the remnants of the Teleut Tele tribes escaping to the then-Russian
Altai fleeing form the army of the Manchurian-ruled Chinese Empire Qing exterminating Jungars and
their former kishtyms without any exceptions (events of 1750s - 1790s), bringing nearly complete extermination
of the Tele people, when no more than 10% of the Tele-confederated people surviving
and finding refuge in the mountainous areas in the Altai, Sayans, Pamir, and Chinese Turkestan. |
Kumans (Kumandins)
Rather ancient branch of the 6 haplotypes (at 7 o'clock) [27 mutations]
13 25 15 10 11 14/15 10 13 11 17/18 - 15 14 20 12 16 11 23 (3800 years to a common ancestor)
(No 8, 51, 59, 64, 65, 72)
This may well be, with the base haplotype in the west of the E.European Plain:
13 25 16 11 11 14 10 13 11 17 - 15 14 20 12 16 11 23
|
This is an allusion to the “Indo-European” Andronovo presence, which is well-established as long
as it is within quotation marks, and to the linguistically “Indo-European” nature of the Andronovo
Kurganians, which is a conflicted concept with far-flung consequences. The ethnological assessment
strikingly conflicts with the linguistic assertions. Ethnologically, the Ural/Siberia migrants,
anthropologically defined as South Eurasian Anthropological Phenotype, and the “Indo-European” aka
“Aryan” southeast migrants were completely incompatible, with the main aspect being in their
relationship to the Kurgan Culture: the aka “Aryan” southeast migrants had none of it, as defined by
the compendium of the archeological traits, plus the aka “Aryans” had agricultural vs. Kurgan animal
husbandry economies, plus archeologically inferred organization of society, plus archeologically and
anthropologically established marital relationships, plus aka “Aryan” religious ideology vs.
etiology expressed in the kurgan tradition, plus markedly different phenotype constitution, plus
opposing lactose tolerance. It is not inconceivable that such disparate societies would speak the
same language, but the only asserted argument for that is the male Y-DNA, while not only the
mother's language is carried over by the female mt-DNA, but also the same Y-DNA studies assert that
Kurgan intrusions into Europe were linguistically non-Indo-European. Even unstoppable Mallory, who
sees Indo-European in every Caucasoid, found it extraordinary difficult to connect Andronovo with Aryan
migration to the northern India. He might find it even less acceptable that he himself
belongs to the “Erbin” stem, converted to the “Indo-Europeism” only lately in the 19th c. If further research would confirm the L342.2 subclade in the Altai
and in the Andronovo culture zone, it would necessitate a re-assessment of the L342.2 lineage
in respect to its presumed linguistic, biological, and ethnological connections. |
Probably, Tele
Young branch of 10 haplotypes at the bottom of the tree, with only four mutations:
13 25 16 11 11 14 10 14 11 18 – 15 14 21 12 17 11 23 (300 years to a common ancestor)
(No 25, 26, 34 - 41)
|
This could be the same Tele migration of 1750s - 1790s, with a different founder |
Probably, Tele
Branch of the seven haplotypes in the upper right part of the tree, in which only five mutations
that gives 5/7/0.034 = 21 generations, or about 525 years to a common ancestor:
13 26
16 10
11 17 11 14
11 18
– 15 14 19 11 15
11 23 (No 43-45, 47-49, 53)
The branch differs by 11 mutations in 17 markers from the base haplotype of the subclades L342.2 (noted above),
13 25 16 11 11 14 10 13 11 17 - 15 14 20 12 16 11 23
that separates their common ancestors by 11/0.034 = 324 → 472 generations, or 11.800 years, and places
a common ancestor of L342.2 and Altai branch at (11800 + 4900 + 525)/2 = 8600 ybp. This common ancestor lived
together with the base haplotype of the E.European Plain 7700 ybp. Again, these are the
autochthonous common
ancestors of the haplotypes. In this (and the next) branch a
characteristic marker is DYS385 = 11-16, while in the E.European Plain it is 11-14.
1523
Probably, Tele
The sub-branch of 9 haplotypes at 3 o'clock (with only 7 mutations,
i.e. the common ancestor lived
7/9/0.034 = 23 generations, or 575 ybp), with a base haplotype
13 26 16 11
11 17 11 14 11 17
– 15 14 19 11 15 11 23 (No 1, 2, 9-11, 12, 15, 52, 54)
It is
apparent that this branch is kindred with the previous branch. They have the same value of DYD385 = 11-17,
and they differ only by two mutations, i.e. their common ancestors diverge by 2/0.034 = 59 → 63
generations, or 1575 years. Their common ancestor lived (1575+525+575)/2 = 1340 ybp. It is clear that this double branch
is young (in respect to their common ancestor). It is visible how
fragmented branches are, how they scatter into recent survivors and equally recent descendants.
|
The timing and scatter resembles the problems at the end of the Western Türkic Kaganate (ca 660s),
whose Kagan Chjenchu was killed, and whose elite had to flee and hide, naturally in the Altai, from
the mobile nomadic armies assembled by the Tang ruler. The modern carriers would tentatively be the
descendents of the Türks/Ashina Türks, a second-largest component of the indigenous Altai-kiji on
the last of their numerous retreats to the Altai. But among numerous Türkic tribes of the time, not
only the Ashina Türks faced problems of survival, and each one of them is a potential candidate for
the fossil DNA. |
A pair of haplotypes (13, 14) are the same, so technically their common ancestor has no age:
13 25 16 11 11 18 11 14 11 17 – 15 14 19 10 15 11 23 (0 years) (No 13, 14)
Kangars/Turgeshes, modern Tubalars and Chelkans
Finally, the last branch (at
5 o'clock) of 6 haplotypes, with a total of 11 mutations, i.e.
11/6/0.034 = 54 → 57 generations, or 425 years to a common ancestor, has a base haplotype
13 24/25 16 11 11 14 11 14 11 17 – 16 14 20 14 16 11 23
(63, 66, 67, 69, 73, 75)
It
sharply differ from the neighboring double branch higher up the tree, especially by GATAH4 (marked).
On the overall, the difference from the base haplotype of the E.European Plain is 6.5 mutations
13 25 16 11 11 14 10 13 11 17 - 15 14 20 11 16 11 23
and from the branch higher up the tree is a 12.5 mutations
13 26 16 10 11 17 11 14 11 18 – 15 14 19 11 15 11 23
producing 12.5/0.034 = 368 → 570 generations, or 14,250 years to the common ancestors. THEIR
common ancestor lived (14250+ 525+425)/2 = 7600 ybp. This is again an autochthonous common
ancestor.
Compiling basic haplotypes for all ten branches of the tree, including the (phantom)
base haplotype of the whole tree at the top (the last haplotype in the matrix), we obtain the following matrix:
1524
13 24 16 9 12
14 10 14 11 18 – 14 14 20 12 17 11 23
13 25.5 16 10
11 14 10 14 11 18 – 15 14 21 12 16 11 23
13 25 16 11 11 14 10 14 11 18 – 15 14 21 10 16 11 23
13 25 15 10 11 14.5
10 13 11 17.5 – 15 14 20 12 16 11 23
13 25 16 11 11 14 10 14 11 18 – 15 14 21 12 17
11 23
13 26 16 10
11 17 11 14 11 18 – 15 14 19 11 15 11 23
13 26 16 11 11 17 11
14 11 17 – 15 14 19 11 15 11 23
13 24.5 16 11 11 14 11
14 11 17 – 16 14 20 14 16 11 23
13 25 16 11 11 18 11 14 11 17
– 15 14 19 10 15 11 23
13 25 16 11 11 14 10 14 11 18 – 15 14 21 12 16 11 23 |
(950 years)
(750 years)
(325 years)
(3800 years)
(300 years)
(525 years)
(575 years)
(425 years)
(0 years)
(0 years) |
Kangars/Turgeshes, modern Tubalars and Chelkans
Potentially Türks/Ashina Türks, or their parent tribe of Saka
Tentatively Tele
Kumans (Kumandins)
Tentatively Tele
Tentatively Tele
Tentatively Tele
Tentatively Tele
Tentatively Tele
Potentially Türks/Ashina Türks, or their parent tribe of Saka |
The whole matrix contains 51 mutations (marked), which produces the age of the common ancestor to 51/10/0.034
= 150 → 176 generations, or 4,400 years prior to the average age of all base haplotypes in the matrix (765 years),
i.e. the common ancestor of all branches lived 4400+765 = 5165 ybp. This may
correspond to the age of the Afanasiev culture (Southern Siberia Nomadic
pastoralism, Southern Eurasian Anthropological Phenotype, 5700/5300 - 4500/4000 ybp, 3700/3300 -
2500/2000 BC).
Literature
Dulik, M.C., Zhadanov, S. I., Osipova, L.P., Askapuli, A., Gau, L., Gokcumen, O., Rubinstein, S., Schurr, T.G. (2012) Mitochondrial DNA and Y chromosome variation provides evidence for a recent common ancestry between Native Americans and indigenous Altaians.
Amer. J. Human Genetics, 90, 1-18. DOI 10.1016/ajhg.2011.12.014
Gonzalez-Ruiz, M., Santos, C., Jordana, X., Simon, M., Lalueza-Fox, C., Gigli, E., Aluja, M., Malgosa, A. (2012) Tracing the origin of the East-West population admixture in the Altai region (Central Asia). PLOS One, 7, 1-11. e48904
Klyosov, A.A., Rozhanskii, I.L. (2012) Haplogroup R1a as the Proto Indo- Europeans and the legendary Aryans as witnessed by the DNA of their current descendants. Adv. Anthropol. 2, No. 1, 1-13.
Klyosov, A.A. (2009) DNA Genealogy, mutation rates, and some historical evidences written in Y-chromosome. I. Basic principles and the method. J. Genetic Genealogy, 5, 186-216.
Klyosov, A.A. (2009) DNA Genealogy, mutation rates, and some historical evidences written in Y-chromosome. II. Walking the map. J. Genetic Genealogy, 5, 217-256.
Klyosov, A.A. (2009) A comment on the paper: Extended Y chromosome haplotypes resolve multiple and unique lineages of the Jewish priesthood.
Human Genet. 126, 719-724.
1525


 tamga, Bulgars (without
Tokhars), Horesmian Ottokars (As-Tokhars), Kipchak Toks-oba of the N.Pontic, Dügers and Digors on
both sides of the Caspian, and few more fragments. In the east, As-Tokhars are known as Ashtak (Ch.
Ashide, tamga
tamga, Bulgars (without
Tokhars), Horesmian Ottokars (As-Tokhars), Kipchak Toks-oba of the N.Pontic, Dügers and Digors on
both sides of the Caspian, and few more fragments. In the east, As-Tokhars are known as Ashtak (Ch.
Ashide, tamga